L’apoptosi è un fenomeno finemente regolato che può innescarsi a causa di una grande varietà di stimoli.
L’imput può essere sia extracellulare (rappresentato dal legame di una molecola segnale esogena con un recettore specifico di membrana, tipo il recettore FAS di morte cellulare) che intracellulare (provocato da danni al DNA).
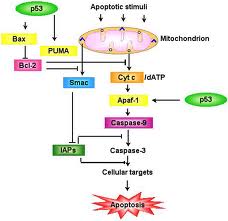
Ambedue le vie di induzione apoptotica, l’extra e l’intracellulare, convergono nell’attivazione di specifici enzimi proteolitici appartenenti alla classe delle caspasi che, di fatto, eseguono il programma di apoptosi.
Nelle cellule molte proteine regolano l’apoptosi, compresa la proteina mitocondriale citocromo C, Apaf-1, i membri delle famiglie Bax, bel-2 e degli inibitori dell’apoptoai (IAP).
Ne deriva un meccanismo comunemente denominato cascata delle caspasi.
Le caspasi una volta attivate, proteolizzano diverse proteine cellulari come le lamine nucleari e inoltre attivano altri enzimi come de DNAasi che dissolvono i cromosomi.
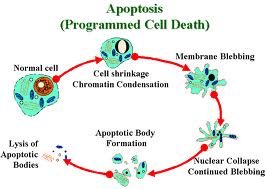
Dal punto di vista microscopico, la prima modificazione morfologica osservabile è la condensazione della cromatina nucleare (picnosi) con formazione di una o più masse compatte vicino alla membrana nucleare.
Contemporaneamente la cellula si stacca dalle cellule adiacenti e il suo citoplasma diventa eosinofilo. Ancora gli organuli citoplasmatici sono indenni.
Con il progredire del processo apoptotico il materiale nucleare si frammenta (carioressi) con successiva disintegrazione della membrana nucleare.
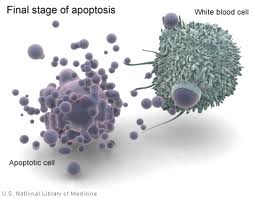
Infine l’intera cellula si rompe (cariolisi) formando frammenti rivestiti da membrana. Alcuni frammenti contengono materiale nucleare e sono noti come corpi apoptotici.
Infine i macrofagi fagocitano i detriti cellulari.








{kind=link}